![G(V, [Ca2+], t) = ˉgm(V, t)pz([Ca2+],t)r, (units: mV, μM, ms)](pub-purkinje-deschutter1-equations10x.png)
Related Documentation:
The channel conductance was determined by the product of voltage-dependent activation (m) and inactivation (h) gates, and for the Ca2+-activated channels a Ca2+-dependent activation gate (z)
|
| (1) |
Equations describing the voltage-dependent gates were described from the classic Hodgkin-Huxley [2] scheme
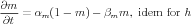 | (2) |
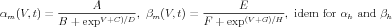 | (3) |
Activation rates for Ca2+-dependent gates were determined by a dissociation constant A and a time constant B
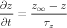 | (4) |
![1
z∞ = -------A---τz = B
1 + [Ca2+]](pub-purkinje-deschutter1-equations14x.png) | (5) |
For the Ca2+ channels the Nernst potential [1] was computed continuously.
Rectification of Ca2+ channels was not modeled using the Goldman-Hodgkin-Katz
(GHK) equation [1] because dendritic membrane potentials in this study stayed
within a range where Ca2+ channels can be considered ohmic (i.e., below -20 mV;
Fig. 4.15 in [1] ). Using the simulation results from the final model, we estimate
that using the GHK equation with an appropriately scaled maximum conductance
( ) to compensate for differences in driving force would cause only small changes
in the amplitude of dendritic Ca2+ currents (mean difference 0.7 %, maximum
4.5 %).
) to compensate for differences in driving force would cause only small changes
in the amplitude of dendritic Ca2+ currents (mean difference 0.7 %, maximum
4.5 %).