Related Documentation:
Ca2+-activated K+ channels are assumed to be responsible for the repolarization of dendritic Ca2+ spikes [13]. Several Ca2+-activated K+ channels have been identified in single channel studies of Purkinje cells [3, 5, 6] among them a large conductance channel corresponding to the BK or maxi-K channel [12]. The macroscopic current carried by this channel is called the C current (KC) and is characterized by a voltage dependence and tetraethylammonium (TEA) sensitivity [1]. This channel is widely distributed in different tissues in both vertebrate and invertebrate preparations, with apparently similar voltage dependence but a variable Ca2+ dependence in all the cells studied [12].
No experimental studies on the kinetics of KC in Purkinje cells were available. Technically it is difficult to characterize the kinetics of KC because the Ca2+ activation cannot be controlled by a “Ca2+ clamp” comparable to voltage clamps. So most experimental investigations have sacrificed temporal resolution by investigating channel activation at steady, well-controlled Ca2+ concentrations [14, 15, 17]. Several groups that have tried to study the temporal dynamics of Ca2+ activation, i.e., how fast the channel reacts to a sudden jump in Ca2+ concentration, have concluded that there was a significant lag in response [4, 8, 9, 11]. Most reports agree that a minimal model of the BK channel requires at least three closed states and one open state, that the open-closed transitions include at least two Ca2+ binding steps and a voltage-independent step, and that the channel does not inactivate [4, 15, 17]. However, there is no agreement on the details of these models because, for example, reported Hill coefficients for Ca2+-dependent opening vary between 1–2 [2, 15], exactly 2 [8, 16] and 3 [9] and some authors assume more than one open state [14, 17]. Most BK channels studied in adult neurons require concentrations of internal Ca2+ in the micromolar range to fully activate [2, 10, 16, 17] and the dependence on Ca2+ concentration seems to be nonlinear [2, 15] (also see however [11]).
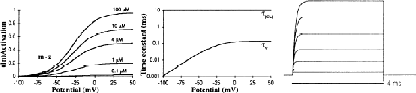
The conflicting experimental data on the BK channel are reflected by the multiple approaches used by different modelers to describe this channel. Most models lump all the open-closed transitions together into one differential equation [7, 15, 18, 20]. Following the example of [19] we have described this channel with two independent state variables (m and z in Eq. 1), but we have used a different model for the Ca2+-dependent step. The Ca2+-independent gate was modeled along data from [4] with a voltage-independent activation (αm) and a voltage-dependent inactivation (βm), with a typical 15 mV per e-fold change in conductance [2, 12]. We shifted the deactivation to more positive potentials to fit the strong depolarizations (>50 mV) required to activate KC in Purkinje cells, as reported by [6]. The Ca2+-binding step was modeled along [2] as an adsorption isotherm distribution with a half-activation at 4 μM and a Hill coefficient of 2 (Eq. 5). The delay in activation was modeled explicitly by a time constant of activation of 10 ms [4, 9, 11].
[1] PR Adams, A Constanti, DA Brown, and RB Clark. Intracellular Ca activates a fast voltage-sensitive K current in vertebrate sympathetic neurons. Nature, 296:746–749, 1982.
[2] F Franciolini. Calcium and voltage dependence of single Ca2+-activated K+ channels from cultured hippocampal neurons of rat. Biochimica et Biophysica Acta, 93:419–427, 1988.
[3] B Gähwiler and I Llano. Sodium and potassium conductances in somatic membranes of rat purkinje cells from organotypic cerebellar cultures. Journal of Physiology (Lond.), 417:105–122, 1989.
[4] M Gola, C Ducreux, and H Changneux. Ca2+-activated K+ current involvement in neuronal functionrevealed by in situ single-channel analysis in helix neurones. Journal of Physiology (Lond.), 420:73–109, 1990.
[5] DL Gruol, VE Dionne, and AJ Yool. Mutliple voltage-sensitive K+ channels regulate dendritic excitability in cerebellar purkinje neurons. Neuroscience Letters, 97:97–102, 1989.
[6] DL Gruol, T Jacquin, and AJ Yool. Single-channel K+ currents recorded from the somatic and dendritic regions of cerebellar purkinje neurons in culture. Journal of Neuroscience, 11:1002–1015, 1991.
[7] M Hines. Efficient computation of branched nerve equations. International Journal of Bio-Medical Computing, 15:69–76, 1984.
[8] AJ Hudspeth and RS Lewis. Kinetic analysis of voltage- and ion-dependent conductances in saccular hair cells of the bull-frog, rana catesbeiana. Journal of Physiology (Lond.), 400:237–274, 1988.
[9] Y Ikemoto, K Ono, A Yoshida, and N Akaike. Delayed activation of large-conductance Ca2+-activated K channels in hippocampal neurons of the rat. Biophysical Journal, 56:207–212, 1989.
[10] B Lancaster, RA Nicoll, and DJ Perkel. Calcium activates two types of potassium channels in rat hippocampal neurons in culture. Journal of Neuroscience, 11:23–30, 1991.
[11] L Landò and RS Zucker. Caged calcium in aplasia pacemaker neurons—characterization of calcium-activated potassium and nonspecific cation channels. Journal of General Physiology, 93:1017–1060, 1989.
[12] R Latorre, A Oberhauser, P Labarca, and O Alvarez. Varieties of calcium-activated potassium channels. Annual Review of Physiology, 51:385–399, 1989.
[13] RR Llinás and M Sugimori. Electrophysiological properties of in vitro Purkinje cell dendrites in mammalian cerebellar slices. Journal of Physiology (Lond.), 305:197–213, 1980.
[14] OB McManus and KL Magleby. Kinetic time constants independent of previous channel activity suggest markov gating for a large conductance ca-activated k channel. Journal of General Physiology, 94:1037–1070, 1898.
[15] E Moczydlowski and R Latorre. Gating kinetics of Ca2+-activated K+ channels from rat muscle incorporated into planar lipid bilayers. evidence for two voltage-dependent. Journal of General Physiology, 82:511–542, 1983.
[16] PH Reinhart, S Chung, and IB Levitan. A family of calcium-dependent potassium channels from rat brain. Neuron, 2:1031–1041, 1989.
[17] TG Smart. Single calcuim-activated potassium channels recorded from cultured rat sympathetic neurones. Journal of Physiology (Lond.), 389:337–360, 1987.
[18] RD Traub. Simulation of intrinsic bursting in CA3 hippocampal neurons. Neuroscience, 7:1233–1242, 1982.
[19] RD Traub, RKS Wong, R Miles, and H Michelson. A model of a CA3 hippocampal pyramidal neuron incorporating voltage-clamp data on intrinsic conductances. Journal of Neurophysiology, 66:635–650, 1991.
[20] Koch C Yamada W and Adams P R. Multiple channels and calcium dynamics. In Koch C and Segev I, editors, From Synapses to Networks, pages 97–133. MA: MIT Press, 1989.